Bau und Funktion der Wurzel


Die Wurzel (Radix)
Wurzeln verankern Pflanzen im Boden, nehmen Wasser und Mineralsalze daraus auf und sind gut geeignet zum Speichern von organischen Stoffen
(Speicherwurzeln). Darüber hinaus werden in der Wurzel Pflanzenhormone (Phytohormone) gebildet und bis in die oberen Pflanzenteile transportiert. Die Ankerfunktion der Wurzel bedingt die Ortsfestigkeit der Pflanzen, die dementsprechend Auswirkungen auf den Konkurrenzkampf hat.
Wurzeln besitzen keine Blätter und somit auch keine Spaltöffnungen. Ihr Leitungsgewebe liegt gebündelt im Zentralzylinder. So sind z. B. Zwiebeln und Knollen (Kartoffel) keine Speicherwurzeln sondern zu Speicherzwecken umgewandelte Blätter und Sprossachsen.
Lässt man Samen (Bohnen, Getreide) keimen, dann bildet sich zuerst die Keimwurzel aus, um den Keimling fest im Boden zu verankern. Erst später beginnen sich die Meristemzellen des Sprosses zu teilen. Die Keimwurzel wächst in die Länge und man erkennt dicht an der Wurzelspitze dünne durchsichtige Härchen, die Wurzelhaare. Sie bestehen aus langgestreckten Zellen der Wurzelepidermis und dienen der sofortigen Mineralsalz- und Wasseraufnahme. Durch viele Verzweigungen und Wurzelhaare wird eine Oberflächenvergrößerung erreicht, die eine höhere Wasser- und Mineralsalzaufnahme gewährleistet.
Das Wurzelwachstum findet zunächst an der Wurzelspitze, am sogenannten Vegetationskegel, statt. Er besteht aus teilungsfähigen Zellen und wird von den abgestorbenen Zellen der Wurzelhaube (Calyptra) und einer Schleimhülle beim Durchstoßen des Erdreiches geschützt. Hinter der Zellteilungszone entsteht später die Zellstreckzone und dahinter findet dann die Differenzierung der einzelnen Zellen zu Gewebe statt. Im älteren Bereich hinter der Wurzelhaarzone entstehen die Seitenwurzeln. Die Anzahl der ausgebildeten Seitenwurzeln hängt stark von den Bodenverhältnissen ab.
Nach der Differenzierung der Gewebe liegen nachfolgend aufgeführte Primärgewebe vor. Ihre Anordnung unterscheidet die Pflanzen in Einkeimblätttrige (die meisten Gräser, Getreidepflanzen und Palmen) und Zweikeimblätttrige (viele krautige Pflanzen und Bäume).
Von außen umgibt die Wurzelepidermis (Rhizodermis) das Gewebe, eine einschichtige Zellschicht, von welcher später einige Zellen die Wurzelhaare ausstülpen. Sie ist somit das wichtigste Absorptionsgewebe der Pflanze. Unmittelbar nach innen angrenzend befindet sich die Wurzelrinde. Ihre äußerste Schicht -die Exodermis- bildet nach Absterben der kurzlebigen Rhizodermis das sekundäre äußere Abschlussgewebe der Wurzel. Zwischen ihr und der innersten Zellschicht der Wurzelrinde -der Endodermis- liegen Parenchymzellen (Rindengewebe), welche häufig der Speicherung von Nährstoffen dienen. Ihre Plasmamembran ist aktiv an der Aufnahme von Mineralsalzionen beteiligt. Im Inneren liegen die Gewebe des Zentralzylinders, der durch die Endodermis vom Rindengewebe getrennt ist. Die Endodermis stellt eine selektive Barriere für den Durchgang von Mineralien aus der Bodenlösung in das Xylem des Zentralzylinders dar. Hier wird der Durchgang der Mineralsalzionen kontrolliert. Nicht alle gelangen in den Zentralzylinder .
In der äußeren Schicht des Zentralzylinders liegen die teilungsfähigen Zellen des Perizykels (Pericambium), aus denen die Seitenwurzeln hervorgehen. Im Zentralzylinder befinden sich die Leitungssysteme:
| |
|
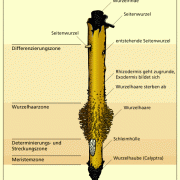
Längsschnitt der Wurzel (Wurzelhaube, Meristemzone, Determinations- und Streckungszone, Differenzierungszone mit Wurzelhaaren)

Querschnitt der Wurzel (Rhizodermis, Exodermis, Rindenparenchym, Zentralzylinder mit Endodermis, Perizykel, Phloem, Xylem, Markparenchym)
Innerhalb des Leitgewebes liegen strahlenförmig angeordnet die Xylembereiche. Zwischen den Xylemstrahlen befinden sich die Phloem-Elemente, welche durch Parenchym getrennt vom Xylem vorliegen. Nach Anzahl der Xylemstrahlen kann man Dicotylen und Monocotylen voneinander unterscheiden. Diarche (2 Strahlen), tetrarche (4 Strahlen) und pentarche (5 Strahlen) Leitbündel finden sich überwiegend bei Dicotylen (Zweikeimblättrige); Monocotylen (Einkeimblättrige) besitzen meist polyarche (vielstrahlige) Leitbündel. Die Strahlenform entsteht bei der Ausdifferenzierung der Leitbündel: Das nach innen gerichtete Xylem entwickelt sich wesentlich stärker und bildet dadurch nach innen gerichtete Strahlen. Ganz selten stoßen diese im Zentrum zusammen (Lauch, Allium), meist befindet sich dort noch das Markparenchym, welches auch verholzen kann (Marksklerenchym).
Wurzeln können durch sekundäres Dickenwachstum, z. B. bei Bäumen, nicht nur in die Länge sondern auch in die Breite wachsen. Dieses Wachstum geht von teilenden Parenchymzellen zwischen den Leitbündeln aus. Es beginnt innerhalb der Phloem-Elemente und setzt sich um die Xylemstrahlen herum fort. Es entsteht allmählich ein Zylinder, die primäre Rinde stirbt ab und wird durch neues (tertiäres) Abschlussgewebe ersetzt.
Die meisten Pflanzen besitzen Wurzeln, die mit Mykorrhiza modifiziert sind. Mykorrhizen sind Pilzgeflechte, die Wurzeln umschließen und tief in die Rindenschicht eindringen. Sie bilden eine Symbiose mit den Pflanzen und erleichtern den Pflanzen die Wasser- und Mineralsalzaufnahme. Der Pilz wird von den Assimilaten der Pflanze ernährt.
Einkeimblättrige Pflanzen (Gräser) bilden nur eine kurzlebige Primärwurzel aus. Die Primärwurzel ist die Wurzel, die unmittelbar aus der Keimwurzel entsteht. Aus ihr gehen später die Seitenwurzeln hervor und bilden ein komplexes Wurzelgeflecht. Bei allorhizer Bewurzelung wird die Keim- bzw. Primärwurzel zur Hauptwurzel, weitere Wurzeln entstehen durch Verzweigung und sind dementsprechend zweit- bzw. drittrangig. Zusätzlich können noch sprossbürtige Wurzeln entstehen.
Bei kurzlebiger Primärwurzel entstehen sprossbürtige Wurzeln als Hauptwurzeln (homorhize Bewurzelung). Alle Wurzeln sind in dem Fall gleichrangig. Sie stellen ein besonders gutes Wurzelsystem gegen Bodenerosion dar. Gebildet werden sie vorwiegend im inneren Rindengewebe, bevorzugt am Bereich der Knoten (Nodien). Man findet diese Art Wurzelbildung besonders bei Monocotylen. Wenn eine Wurzelbildung nach Verletzung an Sprossen oder Blättern erfolgt, spricht man von Adventivwurzeln.
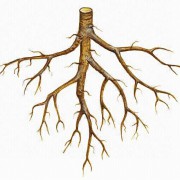
Allorhize Bewurzelung
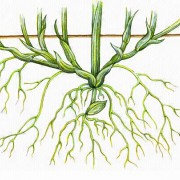
Homorhize Bewurzelung
Einige Pflanzen (z. B. Eichen, Kiefern, Löwenzahn) bilden tief in den Boden ragende Pfahlwurzeln und Seitenwurzeln aus. Es handelt sich hier um eine allorhize Bewurzelung, bei welcher die Hauptwurzel im Wachstum stark gefördert wird und eine tiefreichende Pfahlwurzel bildet (Tiefwurzler). Die Pfahlwurzeln können enorme Tiefen erreichen. Beim Ausbaggern des Suezkanals in Ägypten fand man Tamarisken und Akazien, die Wurzeln bis in 30 m Tiefe ausgebildet hatten. Dagegen erreichen Wurzeln vom Mais nur 1,5 m Tiefe, dafür jedoch seitliche Ausdehnungen von bis zu 1 m. Am genauesten wurde die Wurzel der Roggenpflanze untersucht. Ihre Wurzelfläche betrug 639 m² und war 130 mal größer als das Sprosssystem. Das gesamte Wurzelsystem war in 6 l Erde unterzubringen. Tiefwurzler findet man v. a. in Gebieten mit weniger Wasserverfügbarkeit. Diese ist sowohl Boden als auch Vegetationszonen abhängig.
Bleibt die Hauptwurzel in ihrem Wachstum gegenüber den Seitenwurzeln zurück, spricht man von Flachwurzlern (z. B. Fichte, Pappel). Deren Standfestigkeit ist im Gegensatz zu Tiefwurzlern um einiges geringer, was auch beträchtliche Sturmschäden in Fichtenforsten bewiesen.

Wurzelsysteme
Wurzeln können auch als Speicherorgane ausgebildet werden. Eine Wasserspeicherfunktion übernehmen die Wurzeln von Pflanzen in Trockengebieten der Erde (Xerophyten). Für die Speicherung von Stoffwechselprodukten der Fotosynthese in der Wurzel gibt es die beiden Formen Rübe und Wurzelknolle. Rüben entstehen durch die Verdickung (Dickenwachstum) der Hauptwurzel und treten nur bei Dicotylen auf. An der Rübenbildung können auch Teile der Sprossachse beteiligt sein. Reine Wurzelrüben findet man beispielsweise bei der Zuckerrübe und der Möhre. Bei Rettich, Sellerie und Futterrübe sind sowohl Wurzel- als auch Sprossabschnitte an der Rübenbildung beteiligt.
Wurzelknollen entstehen durch die Verdickung von oft sprossbürtigen Seitenwurzeln. Ihr Auftreten reicht von Monocotylen (Orchideen, Lilien) bis zu Dicotylen (Dahlie, Batate). Die Verdickung und damit Bildung des zusätzlichen Speichergewebes entsteht hier nicht durch sekundäres Dickenwachstum sondern häufiger durch Verdickung der primären Rinde. Verzweigte Wurzelknollen gibt es z. B. bei Orchideen.
In Anpassung an unterschiedliche Lebensbedingungen existieren vielgestaltige Abwandlungen von Wurzeln, die z. T. Funktionen anderer Pflanzenabschnitte übernehmen. Zur optimalen Ausnutzung der (auch ungünstigen) Lichtverhältnisse bilden Kletterpflanzen spezielle Wurzeln aus, die ihnen beim Näherbringen ihrer Fotosyntheseorgane an das Licht helfen. Wurzelranken und Haftwurzeln dienen der Festheftung von Sprossachsen an eine Unterlage.
Bei unzureichender Verankerung von Pflanzen im Boden übernehmen spezielle Wurzel-Metamorphosen die Stabilisierung. Bei diesen handelt es sich um Brettwurzeln, Stelzwurzeln, Stützwurzeln und Zugwurzeln. Häufig findet man diese besonderen Wurzeln bei tropischen Bäumen, die aufgrund ihrer immensen Stammhöhe zusätzliche Stabilität benötigen. Eine besondere Anpassung zeigen einige Orchideenarten. Ihre Vertreter bilden Luftwurzeln, die eine Wasseraufnahme aus der Luft (Epiphyten = bei anderen Pflanzen aufsitzend) ermöglichen. Die Aufnahme des Wassers erfolgt durch tote Zellen eines mehrschichtigen Abschlussgewebes (Epidermis). Das Gewebe besitzt eine schwammartige Beschaffenheit und kann so größere Mengen an Wasser aufnehmen und in die Wurzelrinde weiterleiten.
Zum Schutz vor Fressfeinden bilden manche Pflanzen Wurzeldornen aus. Sie entstehen durch die Verholzung von Wurzelgewebe. Man findet sie z. B. bei einigen Palmen (Acanthorhiza).
Ein Angebot von ![]()