- Lexikon
- Biologie Abitur
- 2 Grundbausteine des Lebens
- 2.4 Zellen und Zellbestandteile
- 2.4.2 Membranen grenzen ab und schaffen Räume
- Membranmodelle im Wandel der Zeit
Membranmodelle im Wandel der Zeit


Im 19. Jahrhundert konzentrierten sich Forschungsarbeiten in Botanik und Landwirtschaft auf die mineralische Ernährung der Pflanzen. JUSTUS VON LIEBIG (1803–1873) hatte um 1840 die Mineraltheorie der Pflanzenernährung begründet und damit direkt und indirekt Anstöße für physiologische Untersuchungen zur Stoffaufnahme von pflanzlichen Zellen gegeben.
Parallel zu den Untersuchungen über die Pflanzenernährung entwickelten sich besonders in der zweiten Hälfte des 19. Jahrhunderts die Forschungsarbeiten auf dem Gebiet des Wasserhaushalts der Pflanzen, dessen Kenntnis auf der Entdeckung der Osmose durch HENRI JOACHIM DUTROCHET (1776–1847) 1826 beruhte. Grenzschichten in Form von Membranen mussten bei diesen Prozessen eine entscheidende Rolle spielen.
In den 60er- und 70er-Jahren wurde die Erscheinung der Diosmose entdeckt, die Semipermeabilität (Halbdurchlässigkeit) der Membran erkannt und ihre Bedeutung für den Stoffwechsel begriffen. Plasmolyse-Versuche zeigten eindrucksvoll Stoffaustauschvorgänge durch biologische Membranen. MORITZ TRAUBE (1826–1894) konstruierte 1867 eine „künstliche Zelle“ (traubesche Zelle) mit einer chemischen, halbdurchlässigen Membran. Dieses Modellprojekt nutzte er zum Studium der Semipermeabilität. WILHELM FRIEDRICH PHILIPP PFEFFER (1845–1920) entwickelte in Leipzig Methoden zur Bestimmung des osmotischen Drucks. Er trug durch seine Osmoseforschungen mit semipermeablen Membranen wesentlich zur Schaffung allgemeiner Grundlagen für die Pflanzenphysiologie bei (pfeffersche Zelle).
CHARLES ERNEST OVERTON (1865–1933) hatte beobachtet, dass nicht nur Wasser und sehr kleine Moleküle bzw. Ionen die Zellmembran passieren konnten, sondern auch fettlösliche Substanzen. Das veranlasste ihn 1895 zu der Feststellung, dass Membranen aus Lipiden bestehen müssten. Die zwei Jahrzehnte später mit Membranen aus Erythrocyten durchgeführten Untersuchungen bestätigten diese Behauptung.
Zusätzlich zu den Lipiden konnte man beachtliche Mengen an Proteinen nachweisen. Im Jahr 1917 stellte IRVING LANGMUIR (1881–1957) künstliche Membranen aus Phospholipiden her. Dafür löste er Phospholipide in Benzol und gab anschließend das Gemisch in Wasser. Nach dem Verdunsten des Benzols fand er als Rückstand einen dünnen Film Phospholipide auf der Wasseroberfläche. Er beobachtete dabei, dass nur die hydrophilen Köpfe der Phospholipide in Wasser eingetaucht waren.
1925 postulierten E. GORTER und F. GRENDEL, Zellmembranen bestünden aus Phospholipiddoppelschichten. Eine solche Doppelschicht könnte eine stabile Abgrenzung zwischen zwei wässrigen Kompartimenten darstellen, weil die hydrophoben Schwänze der Phospholipide – durch die Molekülanordnung – gegen das Wasser abgeschirmt sind, während die hydrophilen Köpfe freiliegen. Dafür maßen GORTER und GRENDEL den Phospholipidgehalt der aus roten Blutzellen isolierten Membran und stellten fest, dass er gerade ausreichte, um die Zelle mit zwei Molekülschichten zu umgeben.
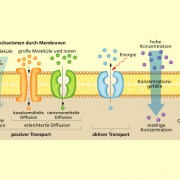
Transportmechanismen durch Membranen
Zehn Jahre später (1935) schlugen J. DANIELLI und H. DAVSON ein Membranmodell vor, bei dem auf eine Phospholipiddoppelschicht beidseitig eine starre Schicht globulärer Proteine aufgelagert ist („Sandwich“). Die von DANIELLI und DAVSON entwickelten Modellvorstellungen haben die Membranforschung in den folgenden Jahrzehnten außerordentlich beflügelt. Immer wieder wurden neue experimentelle Ergebnisse an diesem Modell gewertet und überprüft.
Mit der Entwicklung der Elektronenmikroskopie wurde deutlich, dass Membranen in Zellen weitverbreitet sind und Biomembranen grundsätzliche Strukturen in ihnen darstellen. Nach Fixierung mit Osmiumtetroxid offenbart sich die Membran im elektronenmikroskopischen Bild als dreifache Linie: zwei dunkle, elektronendichte Linien von je 2 nm Dicke getrennt von einer hellen, 3 nm dicken Linie.
J. D. ROBERTSON begründete auf diesen elektronenmikroskopischen Bildern sein Konzept von der Einheitsmembran („unit membrane“). Dieses Konzept widersprach nicht grundsätzlich dem von DANIELLI und DAVSON entwickelten Membranmodell. Dennoch blieben in der Membranforschung viele Fragen offen. Insbesondere zahlreiche physiologische Phänomene konnten nicht umfassend gedeutet werden.
Noch in den 50er-Jahren des letzten Jahrhunderts ist in anerkannten Fach- und Hochschullehrbüchern zu lesen:
„Bei vielen Membranen erfolgt der Durchtritt vorzugsweise durch die wassererfüllten Intermicellarräume, die z. B. in den meisten pflanzlichen Zellulosemembranen so weit sind, dass sie der Mehrzahl der nichtkolloidalen Stoffe keinen nennenswerten Widerstand entgegensetzen. ... Bei anderen können indessen die Poren so eng sein, dass selbst von den echt gelösten Stoffen nur noch kleinere Moleküle hindurchtreten können, während größere Molekülarten wie von einem Sieb zurückgehalten werden. Hierbei ist oft nicht allein der Porendurchmesser, sondern auch die physikalisch-chemische Beschaffenheit der Porenwände (z. B. ihre elektrische Ladung) von Wichtigkeit, da diese die freie Beweglichkeit geladener Teilchen beeinflussen kann.“
Verschiedene elektronenmikroskopische Beobachtungen und Membraneigenschaften ließen sich jedoch auch mit dem bereits modifizierten DANIELLI-DAVSON-Modell nicht ausreichend erklären. Unterschiede im Membranbau, in der chemischen Zusammensetzung und den Flüssigkeitseigenschaften – Fluidität genannt – passten nicht zu den Modellvorstellungen.
Flüssig-Mosaik-Modell
(„fluid mosaic model"; S. J. SINGER und G. L. NICOLSON 1972)
1972 entwickelten S. J. SINGER und G. L. NICOLSON ein Membranmodell, dass mosaikartig angeordnete Proteine auf und in einer flüssigen, in Bewegung befindlichen Phospholipidschicht postuliert. Mit der Bezeichnung Flüssig-Mosaik-Modell (fluid mosaic model) sind die wesentlichen Merkmale dieser Modellvorstellung benannt. Die Phospholipid-Doppelschicht ist in diesem Modell nicht von festen Proteinschichten bedeckt, sondern die Membranproteine sind einzeln in die Phospholipid-Doppelschicht eingelagert und ragen nur mit ihren hydrophilen Bereichen in umgebende wässrige Phase des Cytosols bzw. des Zellaußenmediums. Diese Molekülanordnung gestattet den hydrophilen Regionen von Phospholipiden und Proteinen den größtmöglichen Kontakt mit dem wässrigen Medium und bietet ihren hydrophoben Bereichen gleichzeitig eine nichtwässrige Umgebung. Die Membranen bilden demnach ein Mosaik aus Proteinmolekülen in einer mehr oder weniger flüssigen Doppelschicht aus Phospholipiden. Bis heute lassen sich moderne experimentelle Befunde gut mit diesem Modell erklären. Dies gilt sowohl für die Vielfalt an Membraneigenschaften als auch für die dynamischen Eigenschaften, wie selektive Permeabilität oder Signalaufnahme und Weitergabe. Deshalb wird dieses Modell im Prinzip bis heute anerkannt, einige Modifikationen und Weiterentwicklungen werden jedoch diskutiert.
Modell der abgegrenzten Bezirke
(„fence-and-picket-model"; KUSUMI u. a. 1996)
Proteine sind durch das Cytoskelett an der Membraninnenseite in ihrer freien Beweglichkeit eingeschränkt. Diese abgegrenzten Bezirke können aber übersprungen werden (KUSUMI und Mitarbeiter konnten dies mit einer zeitlich sehr hochauflösenden Kamera und mit winzigen Goldplättchen markierten Proteinmolekülen nachweisen).
Lipid-Floß-Modell
(„lipid-raft-model"; SIMONS u. a. 1997)
Das Lipid-Floß-Modell geht davon aus, dass es in den Membranen floßartige Lipidschollen gibt, die zähflüssiger sind und mit ihren Proteinen in dem Lipidfilm driften. Dabei können einzelne Proteine von diesen Flößen aufgenommen oder abgegeben werden. Die letzten Jahrzehnte brachten eine Fülle neuer Forschungsdaten, aus denen hervorgeht, dass man sich die Membranen eher als Fleckenteppich aus recht unterschiedlichen Regionen vorstellen muss, die sich in Aufbau und Funktion unterscheiden. So weiß man heute, dass die Proteine der Membranen oft zu größeren Komplexen verbunden sind und dass sie nicht – wie im fluid-mosaic-model angenommen – mehr oder weniger zufällig im Lipidfilm driften. Auch die Lipidschicht ist variabel zusammengesetzt. Allein schon die große Zahl verschiedener Membranproteine – bei Escherichia coli konnte man nachweisen, dass das Genom für mehr als 1 000 verschiedene Transmembranproteine codiert – macht eine zufällige Anordnung unmöglich. Dabei haben diese Membranproteine sehr unterschiedliche Formen. Manche sind weitgehend in die Lipid-Doppelschicht eingebettet, andere ragen mit ihren Strukturen weit über die Lipide hinaus und interagieren außerhalb mit anderen Molekülen. Diese herausragenden Strukturen können wesentlich größere Areale überdecken als die in der Lipidschicht steckenden Molekülteile. Außerdem spielen natürlich bei der Membranzusammensetzung auch die verschiedenen Funktionen unterschiedlicher Membranen eine wichtige Rolle. Myelinmembranen enthalten z. B. nur einen geringen Proteinanteil, während fotosynthetisch aktive Membranen sehr proteinreich sind. Viele Membranen grenzen zwei ganz unterschiedliche ,,Phasen" gegeneinander ab, z. B. den extrazellulären Raum gegen das Cytoplasma oder das lnnere eines Zellorganells gegen das Cytoplasma. Diesen unterschiedlichen Milieubedingungen werden die Membranen durch einen ,,asymmetrischen" Bau gerecht: Außenseite und lnnenseite sind unterschiedlich aufgebaut. So sind oft auch besondere Enzymsysteme in Membranen eingebaut, z. B. in den Thylakoidmembranen der Chloroplasten oder in der inneren Mitochondrienmembran.
Funktionen von Membranproteinen
Transport: spezieller Durchlass bestimmter Moleküle oder Molekülsorten teilweise unter Einsatz von Stoffwechselenergie (Hydrolyse von ATP); Enzymaktivität; Signalübertragung; Verbindung verschiedener Zellen; Zell-Zell-Erkennung; Verankerung am Cytoskelett im Zellinneren und an extrazellulären Strukturen, z. B. Kollagenfasern.
Neben Proteinen sind für die Zellerkennung auch membrangebundene Kohlenhydrate von großer Bedeutung (Glykoproteine und Glykolipide).
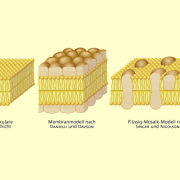
Membranmodelle
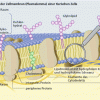
Feinstruktur der Zellmembran einer tierischen Zelle
Membranmodelle im Wandel der Zeit - Feinstruktur der Zellmembran
Renate Diener
Video Irving Langmuir J.D. Robertson DANIELLI-DAVSON-Modell G.L. Nicolson Moritz Traube Biomembranen H. Davson Phospholipidgehalt Membranmodelle Phospholipide E. Gorter Lipidschicht Wilhelm Friedrich Pfeffer Flüssig-Mosaik-Modell F. Danielli S.J. Singer Lipiddoppelschicht Membranen F. Grendel künstliche Membranen Charles Overton Semipermeabilität JUSTUS VON LIEBIG
Stand: 2010
Dieser Text befindet sich in redaktioneller Bearbeitung.
Ein Angebot von ![]()