Fotosynthese


Durch Fotosynthese wird in Pflanzenzellen mit Hilfe von Sonnenenergie Glucose ausund Wasser synthetisiert. Die Kohlenstoffatome des Kohlendioxids werden reduziert und aus dem Wasser wird elementarer Sauerstoff freigesetzt. Im Stoffwechsel der Pflanze kann die gebildete Glucose weiter zu anderen Kohlenhydraten sowie zu Fetten und Proteinen umgebaut werden.
Die Fotosynthese findet in den Chloroplasten statt. Das dort enthaltene Chlorophyll wird durch das einfallende Sonnenlicht angeregt und macht so die Sonnenenergie für die Produktion von Glucose und Sauerstoff verfügbar. Chlorophyll enthält wie das Häm im Hämoglobin ein aus vier verknüpften Pyrrolringen aufgebautes Porphyringerüst. In der Mitte befindet sich ein Magnesiumatom. Die Fotosynthese läuft in zwei eng miteinander verbundenen Abschnitten, den lichtabhängigen Primärprozessen und den lichtunabhängigen Sekundärprozessen ab.
Die lichtabhängige Reaktion besteht im Wesentlichen aus zwei Teilprozessen. Im ersten Schritt wird Licht vom Chlorophyll absorbiert. Das Chlorophyll wird oxidiert und das abgegebene Elektron über mehrere Zwischenstufen auf NADP+ übertragen. Es entsteht NADPH +. Der zweite Teilprozess beinhaltet die Bildung von ATP aus ADP und Phosphat durch den Enzymkomplex ATP-Synthase (Fotophosphorylierung).
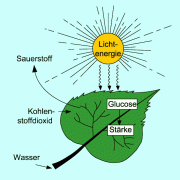
Die Fotosynthese erzeugt Sauerstoff und Stärke und verbraucht Kohlenstoffdioxid und Glucose.
Die ATP-Bildung durch Fotophosphorylierung in den Chloroplasten läuft grundlegend wie die ATP-Bildung in den Mitochondrien während der Zellatmung (Endoxidation) ab (hier Übertragung der chemischen Energie der Nährstoffe auf ATP- oxidative Phosphorylierung). Der Mechanismus der ablaufenden Prozesse wird auch als Chemiosmose bezeichnet.Durch die Aufnahme von Lichtenergie in den Fotosystemen werden Elektronen angeregt und auf ein höheres Energieniveau "gehoben". Damit stehen sie am Anfang einer Elektronentransportkette, die in den Thylakoidmembranen (Bild 2) der Chloroplasten integriert ist. Während die energiereichen Elektronen durch abwechselnde Redoxreaktionen die Elektronentransportkette durchlaufen, werden gleichzeitig Protonen () dem Stroma entzogen. Die Protonen gelangen in den Thylakoidinnenraum und reichern sich dort an. Zusätzlich entstehen bei der am Fotosystem II stattfindenden Fotolyse des Wassers neben Sauerstoff noch Protonen, die ebenfalls die Konzentration an Protonen im Thylakoidinnenraum erhöhen. Die Fotolyse des Wassers findet an der Innenseite der Thylakoidmembranen statt. Dabei wird Wasser durch ein Enzymsystem, mit Mangan-Atomen als aktivem Zentrum, katalytisch oxidiert. Bei der Zersetzung der Wassermoleküle entstehen: Protonen, Sauerstoff und Elektronen:
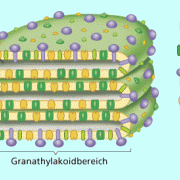
Der Aufbau der Thylakoid-Membran
Die bei dieser Reaktion entstehenden vier Elektronen sorgen für das Auffüllen der Elektronenlücke im Fotosystem II. Gleichzeitig decken die Protonen den Energiebedarf, der für die Reduktion von NADP+ benötigt wird. Dieses Coenzym ist für die Übertragung von Elektronen und Protonen von den Lichtreaktionen auf die Prozesse der lichtunabhängigen Reaktionen notwendig. Der entstandene Sauerstoff ist eigentlich nur ein Abfallprodukt der Fotosynthese. Aber die 2 mal 1011 Tonnen, die dabei jährlich gebildet werden, stellen die Lebensgrundlage für alle aeroben Organismen dar. Die Mangan-Atome im aktiven Zentrum des Enzymsystems sollen dafür zuständig sein, dass sich bei der Fotolyse des Wassers Sauerstoff ohne gefährliche Zwischenprodukte bildet. Es ergaben sich weitere experimentelle Hinweise, dass nicht Wasser, sondern Wasser umwandelndes Kohlenstoffdioxid die Quelle des entstehenden Sauerstoffs ist. Außerdem sollen an der Wasserspaltung noch weitere Ionen beteiligt sein, so z.B. Chlorid-Ionen oder Calcium-Ionen, die für die Beschleunigung der Protonenfreisetzung oder der Funktion des Reaktionszentrums des Fotosystems II verantwortlich sind. Diese Vermutungen der verschiedenen experimentellen Ergebnisse zeigen, dass beim genauen Ablauf der Prozesse noch viele Fragen ungeklärt sind.

Aufbau eines Fotosystems
Während sich der pH-Wert des Thylakoidinnenraumes bei Belichtung in den sauren Bereich verschiebt (pH-Wert 5), wird im Stroma, welches die Thylakoidmembranen umgibt, die Konzentration an Protonen geringer, da Protonen bei der Bildung des Reduktionsmittels NADPH + aus dem Stroma verbraucht werden. So ändert sich bei Belichtung im Stroma der pH-Wert und geht leicht in basische Bereiche (pH-Wert 8). Im Endergebnis der beschriebenen Prozesse entsteht eine hohe Konzentration an Protonen im Thylakoidinnenraum im Vergleich zu einer geringen Konzentration an Protonen im Stroma. Zum Konzentrationsausgleich diffundieren die Protonen aus dem Thylakoidinnenraum durch ATP-Synthase Komplexe entlang des Konzentrationsgefälles zurück ins Stroma, so dass enzymatisch gesteuert ATP durch Fotophosphorylierung von ADP entsteht:
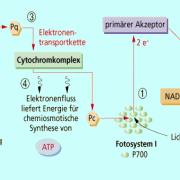
nichtzyklischer Elektronentransport
Die Lichtenergie der Sonnenstrahlen wird von speziellen Antennenpigmenten des Fotosystems I an der Thylakoidmembran der Chloroplasten absorbiert. Die Pigmente sind um ein Reaktionszentrum gruppiert, welches auch als Lichtsammelkomplex bezeichnet wird. Alle Fotosynthesepigmente (Carotinoide, Chlorophyll a, b,c, Phycobiline) verfügen über charakteristische Absorptionsspektren, Hauptpigment der grünen Pflanzen ist das Chlorophyll a - Molekül (Pigment 700, P 700). Jedes Antennenpigment besitzt ein Elektronensystem p, in welchem die Elektronen den Übergang vom Grundzustand in den angeregten Zustand vollziehen. Die Organisation der Pigmente ermöglicht die Übertragung des Anregungszustandes auf ein benachbartes Pigment des Lichtsammelkomplexes. Dieser Prozess wird so lange fortgesetzt bis die Anregungsenergie beim Reaktionszentrum mit dem längstwelligen Absorptionsmaximum ankommt (Chlorophyll a). Dort wird praktisch die aufgenommene Lichtenergie vieler Pigmente gesammelt. Das aktive Chlorophyll a des Reaktionszentrum löst daraufhin die Elektronentransportkette aus: Ein angeregtes Elektron des P 700 gelangt auf ein höheres Energieniveau, wird freigesetzt und von einem Akzeptor aufgefangen. Zum einen können die Elektronen über Ferredoxin auf das Coenzym NADP+ übertragen werden und so zusammen mit den Protonen aus der Fotolyse des Wassers die Bildung von NADPH + H+ (Reduktionsmittel) bewirken. Der Akzeptor NADP+ wird dabei reduziert, indem er die Wasserstoff-Ionen aus der Fotolyse aufnimmt. Im Molekül hinterlassen diese Elektronen somit ein "Loch". Dieses Loch kann von anderen freien Elektronen im Fotosystem II wieder aufgefüllt werden (nichtzyklischer Elektronentransport). Die andere Möglichkeit ist, dass die Elektronen über Ferredoxin, den Cytochrom-Komplex und Plastocyanin an das Reaktionszentrum P 700 wieder zurückkehren und so die entstandene Elektronenlücke selber wieder auffüllen. Dieser Weg, bei welchem die Elektronen an ihren Ursprungsort wieder zurückkehren, wird als zyklischer Elektronentransport beschrieben. Da bei dieser Reaktion nur das Fotosystem I beteiligt ist, findet in dem Zusammenhang keine Fotolyse des Wassers und weder eine Sauerstoffbildung noch NADPH + - Bildung statt. Das einzige Produkt, welches entsteht ist: Adenosintriphosphat (ATP): ADP +P (Photon) ATP. Durch den zyklischen Elektronentransport wird diese Form der ATP-Bildung auch als zyklische Fotophosphorylierung bezeichnet, weil mit Hilfe von Licht/Photonen ADP zu ATP phosphoryliert wird. Diese zusätzliche ATP-Bildung findet vermutlich bei einer genügend hohen NADPH + - Konzentration statt.
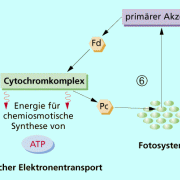
zyklischer Elektronentransport
Das gebildete NADPH und ATP werden in der lichtunabhängigen Reaktion wieder verbraucht. Die einzelnen Schritte der lichtunabhängigen Reaktion wurden von M. Calvin aufgeklärt. Ende der vierziger Jahre verfolgten CALVIN und seine Mitarbeiter wie aus radioaktiv markiertem Kohlenstoffdioxid in einem Kreisprozess Kohlenhydrate hergestellt wurden. Der nach seinem Entdecker MELVIN CALVIN (*1911) benannte Calvin-Zyklus bzw. die lichtunabhängigen Reaktionen (Dunkelreaktion) der Fotosynthese laufen im pigmentlosen Stroma des Chloroplasten ab. Als Voraussetzungen für die Dunkelreaktion dienen ATP als Energiequelle und NADPH + als Reduktionsmittel, die in den lichtabhängigen Reaktionen bereitgestellt worden sind. Licht ist während dieses Prozesses nicht vonnöten. Dabei ist zu beachten, dass sechs Kohlenstoffdioxidmoleküle in den Kreislauf eingeschleust werden müssen, um zwei Moleküle des Primärproduktes Glycerinaldehyd-3-phosphat für eine weitere Verwertung zu Glucose und anderen organischen Stoffen herzustellen.Die ablaufenden chemischen Prozesse und die entstehenden Produkte sind dabei den Vorgängen der Zellatmung ähnlich.Die komplexen Reaktionen des Calvin-Zyklus werden in folgende Teilprozesse unterteilt:
Phase 1: Fixierung von Kohlenstoffdioxid (Carboxylierungsphase):
Den Einbau von Kohlenstoffdioxid in organische Verbindungen bezeichnetman als Kohlenstoffdioxidfixierung. Ribulose-1,5-bisphosphat, ein Zuckermolekül mit fünf Kohlenstoffatomen, welches durch die Anlagerungzweier Phosphatgruppen besonders energiereich ist und als Akzeptor dient, bindet Kohlenstoffdioxid. Die dabei entstehende instabile Zwischenverbindung mit sechs Kohlenstoffatomen im Molekül zerfällt sofort in zwei Moleküle 3-Phosphoglycerinsäure. Dieses Molekül mit drei Kohlenstoff-Atomen ist das erste stabile Produkt im Calvin-Zyklus. Pflanzen, die nur auf diesem Weg Kohlenstoffdioxid binden können, werdenzu den C3-Pflanzen zusammengefasst und sind die am häufigsten Vertretenen unter den Pflanzen. Neben diesem Weg können Fotosynthesespezialisten (C4-Pflanzen, CAM-Pflanzen) in Anpassung an sonnige, trockene Standorte unter günstigen Bedingungen Kohlenstoffdioxiddurch andere Mechanismen vorläufig konzentrieren, bevor sie dann später mit Hilfe des Calvin-Zyklus Kohlenhydrate darmit herstellen. Die Bindung von Kohlenstoffdioxid an den Akzeptor Ribulose-1,5-bisphosphat wird durch eine Carboxylase (Rubisco) gesteuert, welche vermutlich weltweit mengenmäßig das häufigste Enzym ist.
Phase 2: Reduktion:
Bei der Reduktion bindet die 3-Phosphoglycerinsäure eine bei der ATP-Spaltung freiwerdende Phosphatgruppe. Die entstehende 1,3-Bisphosphoglycerinsäure wird durch die Elektronen des NADPH + zu Glycerinaldehyd-3-phosphat (G3P) reduziert, wobei aus der Carboxylgruppe (-COOH) die energiereichere Carbonylgruppe (-CHO) entsteht. Glycerinaldehyd-3-phosphat ist ein Kohlenhydrat mit drei Kohlenstoffatomen (Triose). Zwei Moleküle Glycerinaldehyd-3-phosphat werden aus dem Kreislauf zur weiteren Verwertung entlassen, wobei über mehrere Zwischenprodukte (z.B. Fructose-1,6-bisphosphat) andere organische Stoffe, so weitere Kohlenhydrate (Glucose, Saccharose), Fette und Eiweiße u.a. zur Speicherung von Energie entstehen.
Phase 3: Regeneration des Akzeptors:
Während der Regeneration des Akzeptors entsteht aus den verbleibenden Glycerinaldehyd-3-phosphat-Molekülen zunächst über mehrere Zwischenprodukte (C3-, C4-, C5- und C7 - Verbindungen) Ribulose-5-phosphat. Bevor dann daraus unter Spaltung von ATP und Anlagerung einer Phosphatgruppe derAkzeptor Ribulose-1,5-bisphosphat regeneriert wird.Die lichtunabhängigen Reaktionen lassen sich für die Bildung eines Glucosemoleküls folgendermaßen zusammenfassen:
Die Coenzyme NADP+ und ADP + Pwerden durch die lichtabhängigen Reaktionen wieder regeneriert, so dass sie wieder für den Ablauf des Calvin-Zyklus zur Verfügung stehen.Wenn man alle Prozesse der lichtabhängigen und lichtunabhängigen Reaktionen berücksichtigt, ergeben sich folgende Gesamtgleichungen:
Bruttogleichung:
Nettogleichung:
Dabei wird eine Energiemenge von 2877 kJ verbraucht, um ein Mol Glucose zu bilden.
Fotosynthetisch aktive Pflanzen zeigen einen Gasaustausch. Der Verbrauch von Sauerstoff und die Abgabe von Kohlenstoffdioxid werden formal als Atmung bezeichnet. Dieser Gaswechsel verläuft im Licht wesentlich stärker als im Dunkeln und wird Lichtatmung oder Fotorespiration genannt. Ausgangspunkt für diesen Prozess ist die Doppelfunktion des Enzyms für die Fixierung von Kohlenstoffdioxid an den Akzeptor Ribulose-1,5-bisphosphat. Ribulosebisphosphat-Carboxylase hat insofern eine Doppelfunktion, als es neben der Steuerung der Kohlenstoffdioxidfixierung auch für die Bindung von Sauerstoff verantwortlich ist und als Oxygenase wirkt. Das Konzentrationsverhältnis von Sauerstoff- bzw. Kohlenstoffdioxid entscheidet über den Reaktionsverlauf. Bei hoher Sauerstoff- aber geringer Kohlenstoffdioxidkonzentration wirkt das Enzym als Oxygenase und bringt Sauerstoff in den Calvin-Zyklus ein. Dabei wird der Akzeptor oxidiert und im Gegensatz zur Zellatmung kein ATP hergestellt. Unter natürlichen Bedingungen (20% , 0,03% ) verlieren C3-Pflanzen ca. 50% des assimilierten Kohlenstoffs, der energetisch nutzlos wieder in Kohlenstoffdioxid umgewandelt wird. Die Bedeutung dieses Prozesses ist bis heute unklar. Bei geringem Sauerstoffgehalt im Vergleich zu hoher Kohlenstoffdioxidkonzentration überwiegt die Fixierung von Kohlenstoffdioxid, so wird die Fotorespiration gehemmt und die Nettofotosynthese steigt. Fotosynthesespezialisten (C4- und CAM-Pflanzen) zeigen im Vergleich zu C3-Pflanzen keine Fotorespiration, da das Enzym für die Kohlenstofffixierung (PEP-Carboxylase) keinen Sauerstoff in den Kreislauf einspeisen kann.
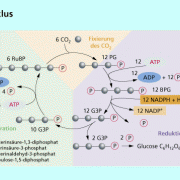
Schema des Calvin-Zyklus
Ein Angebot von ![]()